

How to manage nutrition for better vegetable production
How do we implement the principles of regenerative soil management on a smaller scale — a market farm scale — where we might have 40 or 50 different crops in a half-acre or on five acres, taking into consideration all of the different plants’ nutritional requirements — where we can’t really use tools such as sap analysis like we do on large-scale commercial crops?
This is admittedly a bit of a challenge. I think the foundational framing for this type of approach, though, is for plants to primarily get their nutrition from biology.
Biology supersedes chemistry. What I intend to communicate with that phrase is that you can have soil with perfect chemistry — with mineral nutrition perfectly balanced on a soil analysis —and yet you will not grow healthy plants on that soil as long as you have poor biology. Chemistry alone — without biology — does not grow healthy crops.
This is one of the fundamental failings of hydroponic production. However, the reverse is very possible: when you have soils with vigorous biology, but you have imbalanced mineral chemistry, you can still grow very healthy crops. It’s possible for biology to overcome that, to a degree.

The foundational premise of this entire conversation is that we need to develop robust microbial populations so that we can overcome any potential chemistry imbalances.
Building a robust microbial population, on the surface, seems really simple. And it is, but there’s a lot behind it that it needs to be executed well in order for it to deliver the types of results we’re all looking for.
In summary, first we need to create the right physical environment for soil biology. We need to add the biology that has been lost. We need to constantly feed that biology with mulch or cover crops, and we need to provide balanced mineral nutrition. And possibly, if we’re interested and have the desire to, we can consider speeding up the development of that ecosystem with foliar fertigation applications. And then there are a few other nuances that I want to touch on as well, such as avoiding excessive nutrient applications.
The Rhizophagy Cycle
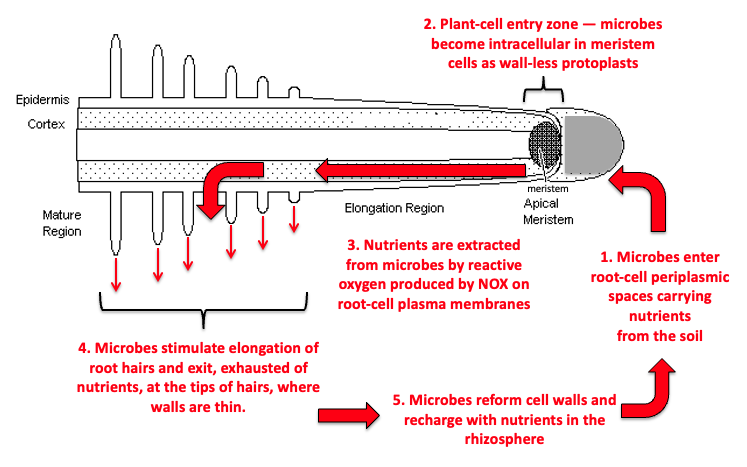
I want to begin by introducing James White’s work on rhizophagy. If you’re not familiar with this yet, you should be, and you will want to learn more about it. The rhizophagy cycle is the idea that plants are getting their nutrition from living bacteria. This is a completely different perspective on plant nutrition than the idea that plants get their nutrition in the form of simple ions — calcium, magnesium, ammonium and nitrate ions — from the soil solution.
As plant root tips extend through the soil, this growing root tip is actually quite porous. It is engulfing — in essence, swallowing or inhaling — entire bacterial cells. These bacterial cells are moving back into the root, and as they move back through the root, and also as the root tip extends, the plant releases nitrous oxide that strips the cell membranes off of these bacteria. So now you have essentially naked bacterial cells — without a cell membrane — moving through the roots of vascular tissue.
Then these naked bacterial cells, or pieces of them, are absorbed by the plant cells through a process known as endocytosis. Plant cells capture these bacterial cells and swallow them whole. And then, once these naked bacterial cells are inside the plant cell, the plant uses the pieces and parts of these bacterial cells — the nucleic acids and different proteins and enzymes — to build its own cell and to maintain that cell.
However, some of these naked bacterial cells survive this process and are not immediately consumed by the plant cells. As they move back through the root system, they trigger the formation of root hairs. And as these root hairs are formed, these naked bacterial cells move back out into the soil environment.
There are a few interesting pieces that happen here. One is that the plant communicates to these bacteria inside the root what its nutritional requirements are — it tells the bacteria, “I need more phosphorus” or “I need more manganese” or “I need more zinc.” Then these bacteria move out through the root hair tips back into the soil environment. And really interestingly, plants do not just release the bacteria back into the soil environment — they also release mucigel and specific carbohydrates and proteins that are precisely what the bacterial cells need to re-form their cell membranes.
The plant is supporting the bacteria specifically so that the bacteria can re-inoculate the soil and then communicate this information to the rest of the soil’s microbial population — to both the fungal and bacterial populations. Then the entire microbial population begins focusing on extracting more of the nutrients the plant needs and providing it to the plant, and the plant continues to feed the microbes. This is a really incredible process. It completely reimagines plant nutrition.
These types of processes have been described in the literature for over 100 years, but they weren’t widely accepted until very recently. This is a really big deal from a plant nutrition perspective and from an agronomy perspective, because for the first time there is an understanding of how plants absorb nutrients that are not simple ions.
We know that there is no one fertilizing the forest; there is no one fertilizing native ecosystems — yet these native ecosystems have adequate or abundant levels of calcium and magnesium and all the different nutrients, including nitrogen and phosphorus and potassium. Where is it coming from? There are only very tiny amounts of soluble ions in the soil solution in native ecosystems, and yet these plants seem to have really good nutrition.
This is an explanation for how plants get nutrients in natural ecosystems, and our domesticated crops — the plants we’re growing for food — have the same capacity. If we want to have robustly healthy plants that have highly functional immune systems and produce high concentrations of plant secondary metabolites, we need them to be absorbing their nutrition in this form of living microbes.
This means that a plant’s digestive system is similar to the rumen of a ruminant animal, or perhaps to our own gut linings, where it is really the biology that provides nutrients to the plant. Nutrition is a biologically mediated process. Plants are farming bacteria in much the same way that we farm livestock. This also means that plants are not vegetarians. They’re consuming entire bacterial cells as a protein source — as an energy source — as their foundational source of nutrition.
Physics: Temperature and Water
What this means from a practical perspective is that we need to consider our topsoil layer as a laboratory petri dish. We want this layer to have abundant and active microbial populations. When we successfully nurture this rigorous biology, it has the capacity to supply the plant’s nutritional requirements, within some boundaries.
It becomes obvious that it is critical to regulate water and temperature. This is important in any environment, but it’s becoming increasingly important given the climactic vagaries that we have experienced in the last decade or two. In today’s environment, it is not enough for us to say, “I’m going to develop really high organic matter soils so that they can hold lots of water, and I’m going to depend on that to mitigate and give resilience during drought conditions and during high rainfall events.” It’s very important to do that — I’m not discounting the need for that — but having high organic matter soils is not enough to mitigate the effects of getting 5 inches of rainfall in 48 hours — or having that happen three times and in a nine-week period.
What does this look like? From a perspective of regulating soil surface temperature, a soil surface should never be bare. When you have bare soil exposed to the sunlight, it can heat up to a temperature of 140 degrees Fahrenheit or warmer on the soil surface and as much as 110 to 120 degrees Fahrenheit 3 to 4 inches down, depending on how the soil was aerated and tilled and how much carbon it has. Enzymes are denatured when you have temperatures warmer than 110 degrees. So anytime we have a bare soil surface exposed to the sunlight, that has the effect of shutting down enzymatic and microbial activity.
If we want to develop these robust microbial populations, we cannot afford to ever have bare soil. Soils need to be covered, either with living plants or with mulch. It’s not acceptable to have soil surface exposed to the sun.
When we think about regulating moisture, our observation and experience has tended to be that microbes can survive dry environments — they go dormant, or they live on the very thin water films on soil particle surfaces or within aggregates. They recover when rainfall reoccurs. But they cannot survive for extended wet periods. An extended wet period is not really all that long — when you have the soil saturated for about 72 hours or longer, that’s enough to deplete all of the dissolved oxygen within the soil aggregates and within the water profile. The moment you’ve depleted all the dissolved oxygen, you have begun changing the soil’s microbial profile and shutting down many of the beneficial organisms that we need to have in these vibrant ecosystems.
So, having saturated soils for an extended period — longer than a day or two — is really not acceptable if we think about managing the soil as a petri dish. I’ve come to the conclusion that given the climate that we live in, drainage is an imperative. There are a few possibilities to make this happen. One is to have a good slope so the surface water moves off quickly, while protecting the soil surface with mulch or cover crops. That might be enough.
But if we don’t have the advantage of a slope, we need to facilitate water moving off quickly in other ways. Over the last decade I’ve observed a new form of field tiling in Northeast Ohio and Pennsylvania that I am incredibly impressed with. Instead of using 4-inch drain tile buried 36 to 48 inches deep and 20 to 25 feet apart, growers are beginning to use shallow drain tile that is only 2 inches in diameter. It’s buried maybe 15 to 18 inches deep, 12 to 15 feet apart.
I have been really impressed with what I’ve observed so far. First of all, the drain tile is shallow enough and close enough together that when we do get those 3-inch or 5-inch rainfall periods, or when we have six or eight weeks of continuous wet weather, they are shallow enough and close enough together to rapidly help that top A horizon — that topsoil layer — to dry out, allowing us to keep our beneficial biological populations. They are also shallow enough that they do not disrupt the natural water table, and they allow the water table to come up high enough that root systems can still reach down into the water table in really dry environments or in drought conditions.
Chemistry: Adjusting the Mineral Profile
Besides adjusting the physical soil profile in terms of managing temperature and managing moisture, the next piece we need to look at is adjusting the mineral profile.
Dr. Elaine Ingham and other microbiologists have said that abundant microbial populations can supply 100 percent of the crop’s nutrition requirements. That can be true, but it’s not necessarily true. It can only be true when soils have the native geology — the native rock profile — that contains all the minerals that the crop requires. We cannot rely on biology to deliver nutrients that are not present.
Our manner of pulling soil analysis and adjusting mineral profiles has shifted over the years. The approach I recommend for market farmers specifically is to pull two different types of soil analyses, and to pull them one time only. The first is a geological assay, also known as a total digestion test. This is essentially a mining assay. Our team has been using Midwest Labs for this type of assay (ask for a PT2 test — Total Minerals Assay — the 2022 price is $22). This type of assay is really valuable because it can show us what minerals are actually present in our soils and our geological profile. The second analysis is a cation exchange capacity (CEC) analysis.
There can be significant differences between these two when we look at different types of parent rock material. One of our consultants had an interesting experience recently. He had geological assays from Midwest Labs side by side from soils that were collected in the Midwest and then also in Florida. On the CEC analysis, both of these soils reported 10 ppm manganese — which is not enough for the Midwest; we need about 20 ppm. For the Florida soil, which is very sandy, the geological assay — the mining assay — also showed that the sum total of what was in that geological profile was 10 ppm manganese. In other words, everything that was there was already showing up on the CEC analysis, so if you wanted to increase manganese in that soil, you had only one option: to add more. However, on the Nebraska soil, the soil also showed 10 ppm on the CEC extraction, but on the geological assay it showed 400 ppm. For that soil, we don’t need to add more manganese — we simply need to develop the biology to extract what is already there.
Biology has the capacity to release these nutrients and make them available for plants — dependent on the geological profile. Use geological assays to determine whether we really truly need to add more, or whether we only need to develop biology in order to release what is already there.
We use the CEC analysis primarily for calcium and magnesium balance, as well as for sulfur, zinc and boron levels — although for zinc we defer to the geological assay to a large degree. For all the other nutrients we defer to the geological assay to determine whether we truly need to add more.
The other factor we need to look at is paramagnetism. We know that plants don’t grow from nutrients alone. Biology doesn’t grow just from carbon, hydrogen, oxygen, nitrogen and so forth. It derives its energy from the energy contained within these molecules.
It’s a bit challenging to talk about energy because it’s such a nebulous concept, and the word “energy” can mean so many different things. But what is quite obvious to anyone who has evaluated the research is that soils that have high levels of paramagnetism support thriving populations of biology, and plants do much better in those environments. I believe it is imperative for us to make sure that our soils have very good paramagnetic levels.
I generally recommend adding paramagnetic rock powders such as basalt — either into compost or onto the soil profile — to rebuild soil paramagnetism. This is somewhat dependent on your native geology. If your native geology is a basalt bedrock, then you probably don’t need to add more.
Biology: Regenerating Activity
Now that we’ve looked at shifting the physical environment and adjusting the mineral nutritional environment, we get to the heart of the conversation of the need to regenerate biological activity.
I would propose that on a market farming scale, we need to build our own high-quality compost. I’ve come to respect and appreciate the results that I’ve observed with the Johnson-Su bioreactor, and it’s something that is appropriate and easy to create on a small scale. You can also use vermicompost. I’m an advocate of doing both. That raw material can then be used to develop compost teas for fertigation and for foliar applications, particularly for disease control.
It can be useful to add any needed soil amendments into the compost, although compost is not the place to add materials like calcium or magnesium for pH corrections. The composting system can easily become too alkaline or too acidic. It’s best to apply those materials directly to the soil. But trace minerals can certainly be added to compost, and this generally has the effect of increasing their bioavailability.
In a market farming context, I highly recommend avoiding the use of animal-manure-based compost or soil amendments. We have observed with our sap analysis work that the majority of insect and disease problems are the result of excess nutrients that are applied by growers — not by deficiencies. The most common excessive nutrients are nitrogen, phosphorus and potash, which are particularly concentrated in animal-based manures. I have observed that it is very difficult to develop excess levels of nitrogen, phosphorus and potash in plant-based compost; as long as there is good biology, we don’t seem to easily develop excesses.
Market farms often apply compost in large quantities as a mulch for weed control — often several inches thick. The quantity of nutrients added is astoundingly high, and it is this overapplication, particularly of nitrogen and potassium, that is at the foundation of most insect challenges — flea beetles, and aphids, and the list goes on. I highly suggest avoiding excesses of those materials.
I also have an unusual suggestion in regard to compost teas. Compost teas, whether they are from Johnson-Su bioreactors or from vermicompost, in my opinion, are not enough to re-inoculate soil biology. They’re very powerful and I’m strongly in favor of them, but many of the really beneficial microorganisms that colonize the rhizosphere require a living plant root to propagate. Mycorrhizal fungi is a classic example — you will not find mycorrhizal fungi in vermicompost. You will not find mycorrhizal fungi in any compost that does not have living plants growing on the compost pile; you have to have living plants and growing root systems to propagate some of this really important biology. For this reason, in our consulting work on larger-scale operations, we use purchased inoculants — both mycorrhizal inoculants and others — because there is a lot of biology that you cannot replicate in a composting system.
Something else I want to point out is that while compost tea applications are very valuable, and they do a lot to regenerate soil biology, they’re not enough to get to where we want to go quickly enough. The next step — once you’ve added biology and the physical environment — is that you need to feed biology every day. The biology needs to eat. We know that the smaller the animal, the more frequent feeding it requires — consider a human baby versus an adult. Imagine what that looks like when you get down to the size of a microbe. They need a constant food source all the time. There cannot be a delay; that cannot be shut off. Biology needs to be fed constantly, either with a mulch or straw or hay or with cover crops.
I’m a big fan of cover crops because they do things that mulch does not. Cover crops stimulate soil aggregation. Our goal is to have that entire A horizon — that topsoil layer — really well aggregated, so you can put your hand in as deep as that topsoil layer goes without any problems. Cover crops have the effect with their root systems of producing soil aggregation particles much faster than mulch does by itself.
| Top Disease-suppressive and Soil-aggregating Cover Crops: – Oats – Buckwheat – Mustard – Tillage radish – Forage rape – Annual ryegrass |
Also consider the concept of using synergistic stacks when you put together foliar applications and fertigation applications. A synergistic stack may combine compost tea, a microbial inoculant like seaweed materials, plant nutrients and trace minerals. You can put all these together.
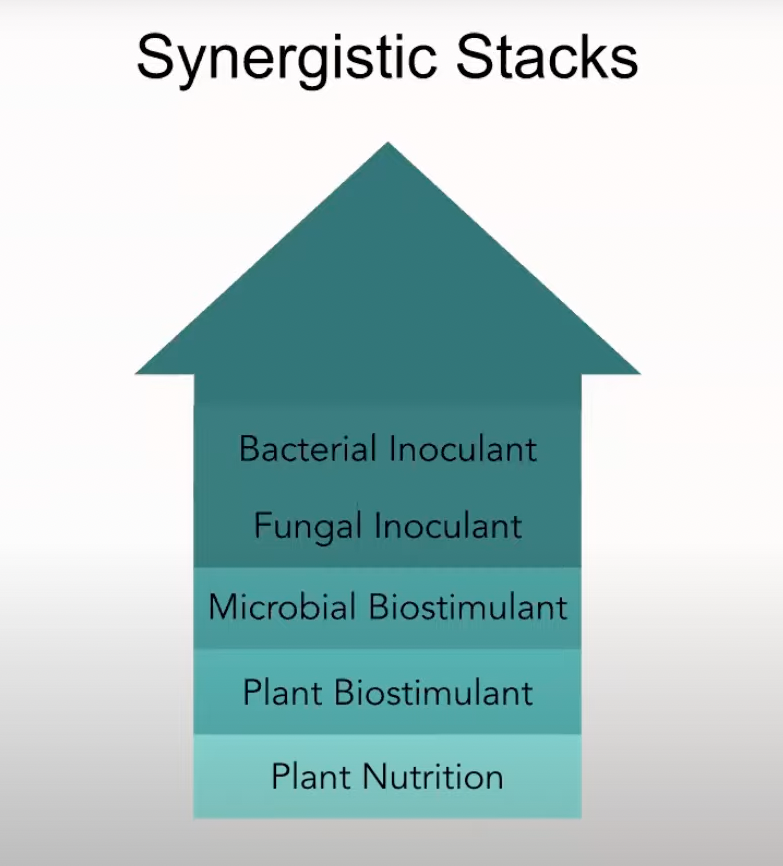
The intention, in terms of speeding up the overall ecosystem, is really photosynthesis, in which the plant captures carbon dioxide and produces sugars and sends more sugars out to the soil biology to reinvigorate and speed up soil biology. The more sugars we can send to the root system, the faster the soil biology will develop. So, increasing photosynthesis is really the pathway to achieving that in the most effective way possible. This is why I’m such a fan of using foliar applications to increase the photosynthetic potential of a crop.
Also, compost teas and microbial inoculants, when used for the long term — particularly if they’re well produced — will have the effect of enhancing disease suppression on the leaf surface. You will have fewer challenges with mildews and rusts on the leaf surface.
In summary, treat your soil as a biological lab. Create the needed physical environment. Add biology that has been lost and feed it with mulch or cover crops. Provide balanced mineral nutrition — but avoid adding excess nutrients — and consider speeding up the system with foliar applications and fertigation.
This sounds so simple. And it really is — it’s not difficult. But you cannot skip steps. You can’t cut out part of the recipe. It takes all these pieces together to produce the really regenerative ecosystem we’re looking for.
John Kempf is the founder of Advancing Eco Agriculture and is the executive editor of Acres U.S.A. magazine. Learn more at johnkempf.com. This article is an edited transcript of a presentation originally given at the 2021 Bionutrient Food Association Conference.
| Have you working to develop healthy soil on your farm? Share your experiences with the Acres U.S.A. community by telling us about it via email (editor@acresusa.com), social media (@acresusa on FaceBook, Instagram, Twitter and LinkedIn) or mail (P.O. Box 1690, Greeley, CO 80632). And sign up for our newsletters at ecofarmingdaily.com to learn lots more about the transition process. |










{kind=link}