
Genetically modified crops affect root exudates and the soil microbial community
Organic substances released into the rhizosphere comprise 20 to 40 percent of the carbon captured through photosynthesis, with the actual amounts depending on the particular plant species. These root exudates are vital nutritional sources for the soil microbial community and are selective for the microbial species that inhabit the rhizosphere and endorhizal micro-environments of the host plant.

Root exudates are low-molecular-weight compounds (organic acids, sugars, phenolics, amino acids, fatty acids) that contain some complex compounds, including sterols, flavonoids, vitamins, nucleic acids, gibberellins, alkaloids, and terpenoids.
The microbiome selected by root exudates may acquire available N, P, K, and S for plant uptake, increase the availability of micronutrients, synthesize antibiotics for protection against pathogens, provide plant-growth-promoting compounds, aggregate soil and organic particles for optimum aeration and water movement, and protect plants from abiotic stresses.
Many factors influence the rhizosphere microbiome, including plant photosynthesis mode (C3 vs C4), plant variety and chemical signaling ability, soil factors (texture, organic matter, pH, etc.), agroecosystem management (tillage, agrochemical use, synthetic fertilizers, manure, etc.), and plant diversity within the host plant rooting zone.
Despite the widespread production of genetically modified (GM) crops, information regarding their influence on biological processes, including root exudation, remains limited. Numerous reports indicate that GM crops affect several soil health and environmental services mediated by environmental microbiomes. This impact is attributed partly to nonspecific or indirect consequences, known as pleiotropy, resulting from the genetic transformation.
For instance, GM soybeans may exhibit excessive exudation of soluble carbohydrates and amino acids from roots, leading to alterations in the rhizosphere microbial community structure compared with non-GM cultivars. This can attract dense colonization of roots by potentially pathogenic fungi (Fusarium) and bacteria (Agrobacterium) while decreasing the abundance of beneficial Pseudomonas, other growth-promoting bacteria, and rhizosphere microorganisms providing available Mn for plant uptake. Any intended or unintended alteration in the composition or quantity of well-known root exudate components (sugars, organic acids, amino acids, etc.) can impact the soil microbiome. Additionally, GM plant root exudates may contain new substances, including toxins.
A classic example of genetic modification that resulted in known, unintended compounds in the plant is the GM version of the White Russet potato. It was developed to provide blemish-free potatoes for French fries in the fast-food industry (see the January 2019 Acres U.S.A. interview with Dr. Cauis Rommens). It was found that genetic modification disrupted the potato’s metabolic pathways, resulting in the accumulation of at least major compounds in the potato tuber that are normally not found, except possibly at trace levels, in non-GM potato. The compounds — chaconine-malonyl, alpha-amino-adipate, and tyramine — are not only new substances in the potato tuber, and we may assume also found in roots, but are toxic to humans.
Furthermore, the GM-potato tubers were rife with fungal propagules of Fusarium and Verticillium, which could be sources of disease outbreaks and mycotoxins. One of the genes silenced in the GM potato mediates assimilation and transport of nitrogen from the soil through roots, suggesting that limited N decreased the release of amino acid and peptide exudates through roots, resulting in altered rhizosphere microbiome composition.
Maize, cotton, rice, soybean, and eggplant have been modified to incorporate cry genes (i.e., Cry1Ab, Cry1Ab/Ac) from Bacillus thuringiensis that code for the synthesis of an insecticidal protein in Bt cultivars that kills lepidopteran pests; however, these insecticides, or cry toxins, are also released into the soil through root exudation and affect soil and rhizosphere microbial diversity.
Root exudates of Bt cotton promoted spore germination and mycelial growth of the F. oxysporum wilt pathogen relative to root exudates from their respective, non-GM parental lines. Additionally, resistance to Verticillium wilt was lower in Bt cotton lines, in which root exudates also promoted the growth of the Verticillium wilt pathogen. More problematic are the effects on beneficial mycorrhizal fungi. Spore germination and root-attaching hyphae of the mycorrhizal fungus Rhizophagus irregularis decreased in Bt cotton compared with the corresponding parental non-GM cultivars; subsequent root colonization intensity, and arbuscule abundance, were lower in Bt roots. The Bt trait significantly contributed to the inhibition of pre-symbiotic development and mycorrhizal colonization, which may be due to either Bt toxicity or interference during signaling between mycorrhizae and host roots.
Carbon metabolic patterns by the soil microbiome were altered in fields planted to Bt cotton, suggesting that the functional diversity of soil microbial communities was affected by Bt cotton. Continuous cultivation of Bt cotton for more than four years resulted in the persistence of Cry proteins in the soil, which contributed to significant reductions in soil microbial biomass carbon, microbial activities, and numerous soil enzyme activities. Apparent inhibition of soil protease activity by Cry proteins likely suppressed the hydrolysis of soil Cry proteins and thereby contributed to their long-term persistence in soils. Thus, potential adverse effects on soil health biological components likely persist under continuous Bt cotton.
Several Bt corn cultivars reduced soil respiration, had poor mycorrhizal root colonization, and altered rhizosphere bacterial community structure. Growth of Bt corn on tropical soils significantly reduced the abundance of ammonia-oxidizing bacteria (nitrifying bacteria). Abundance of protist and nematode communities are reduced in fields cultivated with Bt corn, which disrupts the functionality of the soil food web.
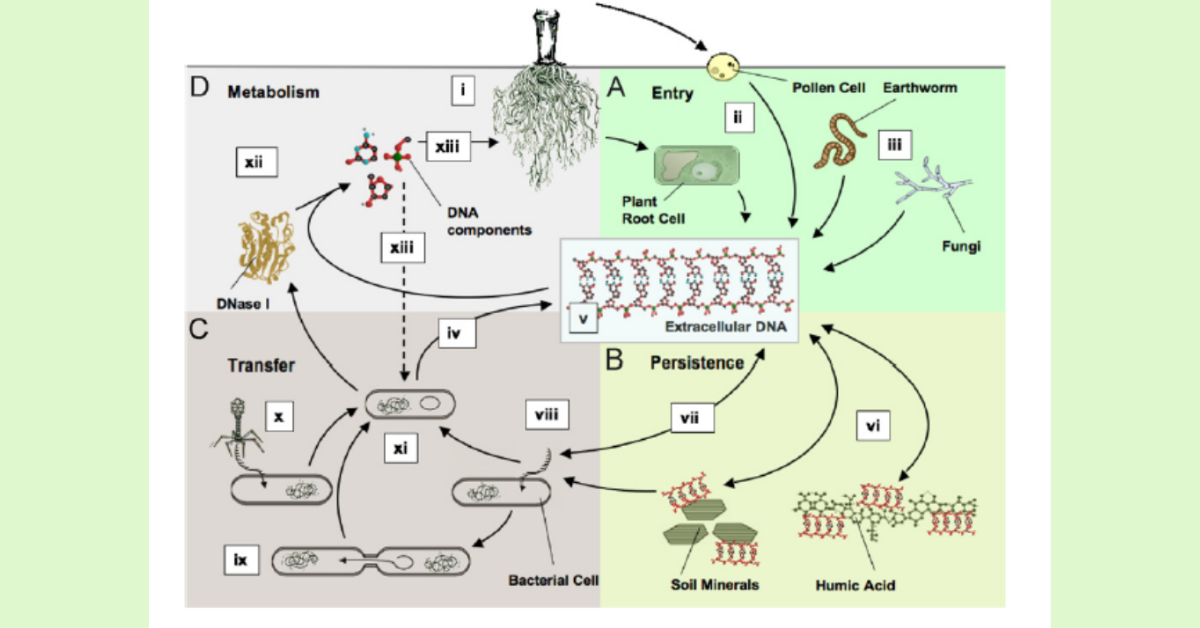
Perhaps far more concerning is the release of intact transgenic DNA through root exudation that can be incorporated by the soil microbiome through horizontal gene transfer. If these genes are incorporated by members of the microbiome into their genome, the resulting intercellular persistence and amplification of these GM DNA sequences within the overall microbial gene transfer network could lead to unforeseen soil biological impacts. Transgene transfer from GM cultivars of transgenic oilseed rape, tobacco, and alfalfa through roots and vegetative residues into rhizosphere soil has been reported to be incorporated into soil bacteria. Implications are that such extracellular DNA released into soil, although at very low concentrations, poses a risk, as it can be a source of the gene pool for neighboring microbial communities, especially bacteria and fungi capable of natural intake of DNA, as well as various phages (viruses) associated within soil bacteria that could incorporate DNA within their genomes and transfer from host to other organisms. Genes coding for Bt toxins within bacteria and fungi could result in suppressing the growth and activity of vulnerable micro- and meso-faunal predators (mites, collembola, other microarthropods) and detrimentally impact nutrient cycling and physical maintenance mediated by the soil food web.
Glyphosate-resistant crop cultivars were developed by the insertion of a transgene (cp4) from an Agrobacterium species to code for an insensitive version of 5-enolpyruvyl-shikimate-3-phosphatase synthase (EPSPS), a critical enzyme required for the synthesis of aromatic amino acids and phenolic compounds important in metabolic reactions, but blocked in glyphosate-sensitive plants and microorganisms. Soil microorganisms are indirectly affected because glyphosate applied to herbicide-resistant crops is released through roots into the rhizosphere environment. Negative effects of glyphosate in the rhizosphere include disruption of legume nodulation by rhizobia and spore germination inhibition of mycorrhizae.
Approximately 80 percent of GM crops under cultivation have modified genes expressing tolerance to glyphosate or glufosinate herbicides and/or stacked with insect resistance. A consequence of widespread use of glyphosate on GM herbicide-tolerant crops is the development of herbicide-resistant (HR) weed infestations that may alter the soil microbiome and soil biological processes that differ from weed-free GM crop monocultures. Depending on the HR weed biotype, infestations may change the amount and type of soluble C released from roots into soil, affect mycorrhizal viability and abundance, and alter plant nutrient uptake in production fields.
For example, in certain rice-growing regions, glyphosate-resistant barnyardgrass releases root exudates that recruit a less diverse bacterial community than the herbicide-susceptible biotype. However, the altered rhizosphere bacterial community functions to aid adaptation of resistant barnyardgrass in the field environment, suggesting that rice tends to be highly vulnerable to competition by the resistant barnyardgrass.
We are learning that several GM crops and trees negatively affect the soil fungal community, even though expressed traits were not expected to affect fungi. This is of concern considering the important roles that fungi perform in soil structure formation, nutrient transfer (including to plants through mycorrhizae), and organic matter formation. Overall, potential impacts on the soil microbiome via altered root exudation during development of GM crop cultivars have seemingly been overlooked, thus emphasizing the need for monitoring key sensitive microbial groups that mediate important soil health processes in future environmental impact assessments of GM crops.
Finally, we need to consider the actual “safety” of newer genetic manipulation methods toward biological processes of root exudation and microbiome response. The genome-editing technique known as “clustered regularly interspaced short palindromic repeats” (CRISPR) allows manipulation of the plant genome at defined sites to allegedly provide more controlled introduction of specific alterations for gene silencing or expression in contrast to random mutations associated with older GM methods. However, recent reports of off-target mutations in base editing of the rice genome using CRISPR suggest that the technique could result in unintended effects expressed in the engineered progeny. The subsequent impact of potentially newly released gene-edited crops on root exudation and soil health relative to older GM crops is yet to be determined.
It is important for farmers to understand the potential soil biological obstacles that arise under continuous production of GM crops in the same fields when considering a transition to non-GM and ecological agricultural production systems. The altered soil microbiome structural and functional diversity, and the persistent presence in soil of DNA and/or products of genetic manipulation, can be remediated by intercropping or integrating mixtures of cover crop species from multiple functional groups (i.e., grass, legume, brassica) that release different organic compounds to stimulate a return to diverse soil microbiomes and restore the necessary beneficial functions of the soil ecosystem.
Dr. Robert Kremer is an adjunct professor of soil microbiology in the School of Natural Resources and the Division of Plant Science & Technology at the University of Missouri and is a retired microbiologist with the USDA Agricultural Research Service.
Information for this article was based on Kremer, R.J. 2020. “Impacts of Genetically Engineered Crops on the Soil Microbiome, Biological Processes, and Ecosystem Services,” in Chaurasia et al. (eds.), GMOs: Topics in Biodiversity and Conservation. Springer.














{kind=link}